
Anche in Italia la raccolta del muschio sta diventando sempre più comune. Questo perché è molto apprezzato nelle composizioni di Terrari e nell’arte del bonsai. Alcune specie rischiano addirittura di scomparire a causa del calpestio dell'uomo, della raccolta eccessiva da parte dei commercianti e, per le specie rare, dell'eccessiva raccolta da parte di alcuni appassionati.
Osserva i seguenti consigli e goditi l'osservazione del muschio:
(1) Non rimuovere tutto il muschio e le piante circostanti.
(2) Raccoglilo solo nelle aree dove è possibile secondo legge. In alcune aree, come terreni privati, parchi naturali e foreste nazionali, la raccolta è severamente vietata dalla legge. Soprattutto nelle foreste vergini, in alta montagna e nelle paludi, si può ritenere che la raccolta sia vietata in tutte le aree.
(3) Goditi la natura.
(4) Fai attenzione a non danneggiarlo!
(5) Osservando la bellezza del muschio nei campi e nelle montagne, posso capire la sensazione di volerlo coltivare in casa. Ma il muschio è bello perché è in natura. Non importa quanto bene lo coltivi, non potrà mai eguagliare la bellezza del muschio naturale.
(6) La nostra missione, da appassionati del muschio, è mantenere intatto l'ambiente naturale in modo da poter osservare un bellissimo muschio ogni volta che visitiamo quel posto. Fai attenzione a non farti distrarre dal muschio che stai cercando, finendo per calpestarne o danneggiarne altro sotto i tuoi piedi o le piante circostanti.
(7) Quando lo raccogli, fai attenzione a ridurre l'impatto sulla comunità di muschio e mantieni la quantità che raccogli al minimo. Anche dei piccoli buchi iniziano a seccarsi e l'intera colonia potrebbe scomparire.
Conosciamo il muschio.
Che cos’è esattamente il muschio? Questa sezione introduce le caratteristiche dei muschi, le loro forme e i vari termini più utilizzati.
Che tipo di pianta è il muschio?
Il muschio è la prima pianta comparsa sulla terraferma, l’equivalente delle prime piante terrestri e si differenzia dalle alghe che vivono nell’acqua. Il muschio cresce ovunque, ai bordi delle strade, nelle foreste, sui dirupi alpini e persino in Antartide. Ovunque andiamo, è sempre lì, come se fosse una cosa ovvia. Per questo motivo tendiamo a dimenticare la meraviglia di questo minuscolo corpo che ha la forza di sopravvivere a siccità, caldo e freddo estremi, cosa che nessun’altra specie vegetale può fare. I muschi non sono irresistibili solo per il loro aspetto, ma anche per il fascino profondo del loro modo di vivere.
Che cos'è il muschio?
Cosa rispondereste se qualcuno vi chiedesse: "Cos'è il muschio? È verde e ama i luoghi umidi. Cresce sulle rocce. Questa risposta, che si sente spesso, è in realtà un'ottima caratterizzazione del muschio. Aggiungiamo quindi un po' di prospettiva scientifica a questa immagine e spieghiamo meglio il muschio:
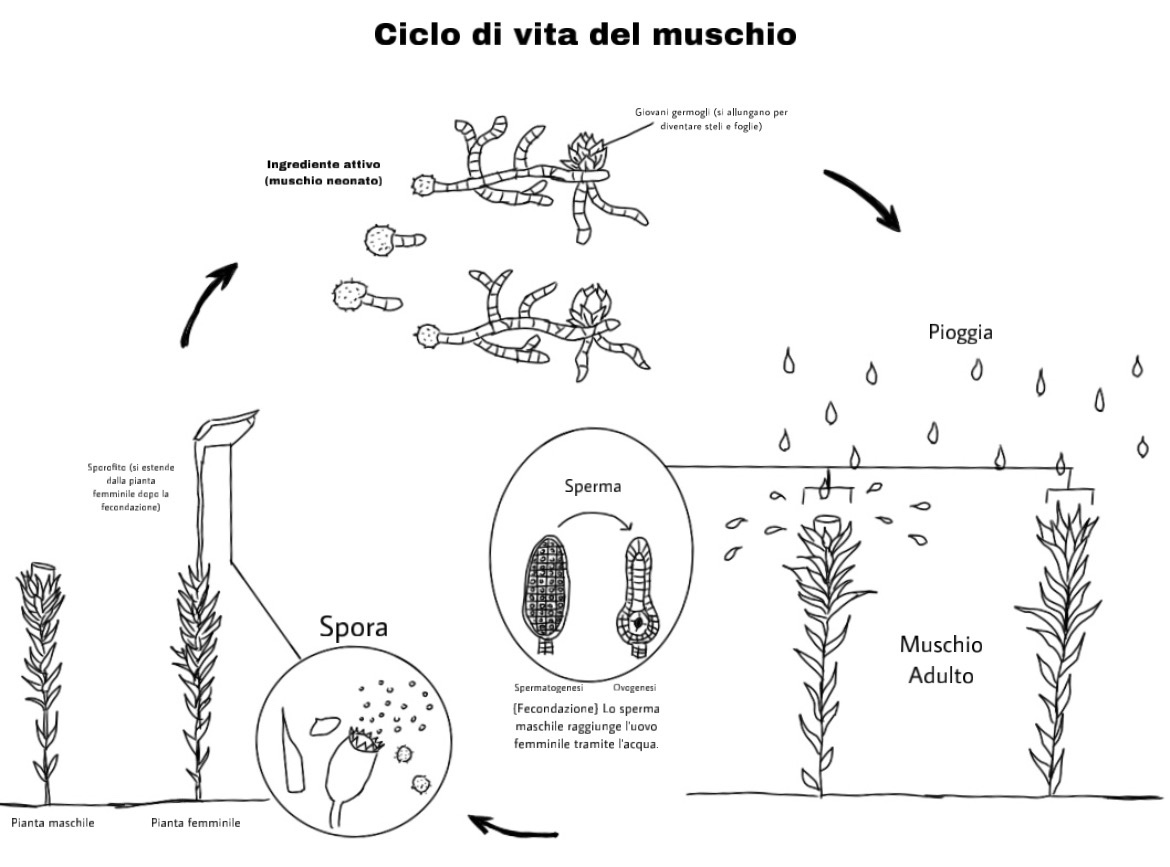
A differenza degli alberi e delle erbe, i muschi non possono produrre “semi grandi e vitali” e non hanno “fasci vascolari” che permettono loro di assorbire l’acqua dal terreno attraverso le radici e distribuirla in tutto il corpo. Si riproducono, invece, per mezzo di spore piccole ma numerose, che permettono loro di trovare rapidamente gli habitat adatti. Inoltre, assorbono la pioggia e la nebbia direttamente dalla superficie delle foglie, consentendo loro di crescere sulle rocce senza terreno. I muschi sono un gruppo di piante che utilizzano la fotosintesi per ottenere energia, ma hanno una struttura corporea molto diversa dalle alghe (alghe verdi) e dalle piante e dagli alberi. Da un punto di vista evolutivo, gli antenati dei muschi vivevano originariamente nell’acqua, ma nel corso del tempo alcune popolazioni hanno sviluppato dei sistemi per proteggere gli individui appena nati, sensibili all’essiccazione, permettendo loro di lasciare l’acqua e vivere sulla terraferma. In sintesi, i muschi sono “piante terrestri che si riproducono per spore e non hanno fasci vascolari“.

Quando si tratta di muschi è meglio avvicinarsi.
Il modo migliore per osservare e riconoscere il muschio è utilizzare una lente d’ingrandimento, al di là della quale vi apparirà un mondo in miniatura tutto da scoprire. Una fitta foresta dove nessun umano ha mai messo piede prima.




Quando ci si addentra nei cespugli verde brillante, si rimane stupiti nel vedere steli e foglie finemente lavorati, forme originali e ricche di particolari. Più si guarda da vicino, più la curiosità viene solleticata. I muschi sono di così piccole dimensioni e così minuscole le differenze tra le specie che per ottenere il nome corretto in alcuni casi è necessario ingrandire al microscopio e osservare la forma dettagliata delle foglie e degli steli, o persino la forma delle cellule. Tuttavia, se vogliamo studiare meglio i muschi, la maggior parte dei principianti senza una lente di ingrandimento non ha altra scelta che tenersi alla larga da questo mondo magico. Sarebbe un peccato sprecare il loro tempo per qualcosa che non potranno apprezzare fino in fondo!
Caratteristiche e struttura corporea dei tre muschi
Esistono tre gruppi di muschi:
- Marchantiophyta (o epatiche)
- Anthocerotophyta (o antocerote)
- Bryophyta (o muschi)
Come primo passo per conoscerli, cerchiamo di capire le caratteristiche di questi tre gruppi.



Sebbene non si sappia ancora con certezza quale dei tre lignaggi sia il progenitore di tutte le altre piante terrestri, sappiamo che la storia più antica delle piante in ambiente terrestre è inestricabilmente legata alla storia delle briofite. Se desideriamo comprendere gli aspetti fondamentali della struttura e della funzione delle piante terrestri, dobbiamo rivolgerci a queste piccole meraviglie della natura.
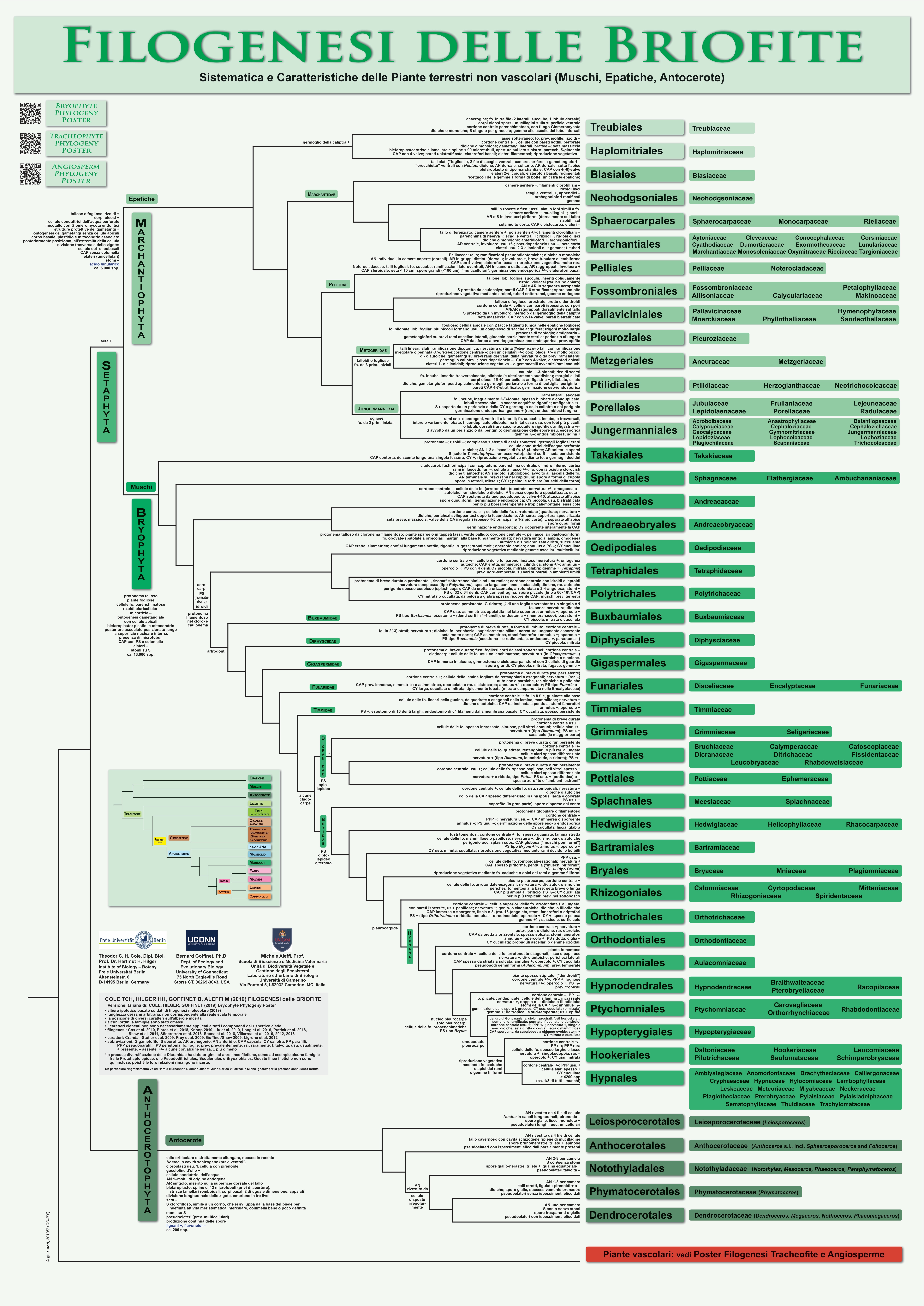
Questi aspetti comprendono:
- l’origine e la natura della crescita tridimensionale delle piante dalle cellule apicali e dai Meristemi .
- l’evoluzione dei meccanismi mitotici cellulari
- lo sviluppo di spore (e poi pollini) spesse, resistenti all’acqua e alla decomposizione
- le pareti molecolari della pianta e del polline
- i meccanismi molecolari e biochimici alla base della tolleranza al disseccamento
- la struttura, la funzione e l’evoluzione del genoma delle piante.
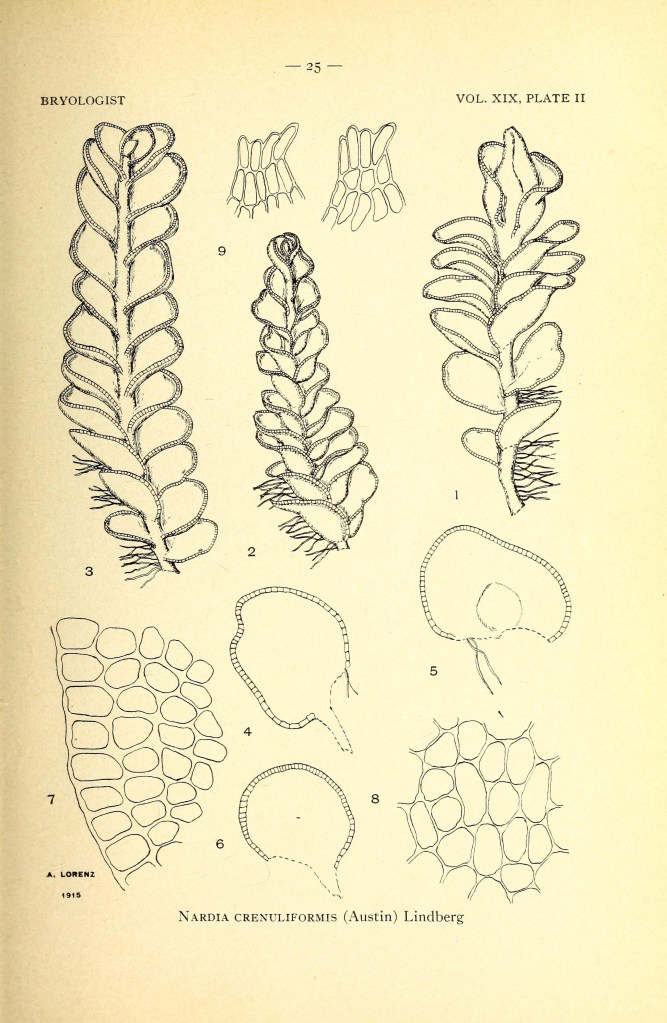
Le Epatiche (Marchantiophyta)
Le epatiche sono un Phylum eterogeneo di piccole piante erbacee terrestri, si stima che comprendano circa 5000 specie in 391 generi.
Occupano un’ampia gamma di habitat, compresi i terreni lungo le rive dei corsi d’acqua, i bordi stradali e i sentieri, nonché le rocce, i tronchi e gli alberi nei paesaggi naturali. Sono presenti in tutti i continenti, compreso l’Antartide, ma sono più diversificati nelle foreste pluviali montane dell’emisfero meridionale.
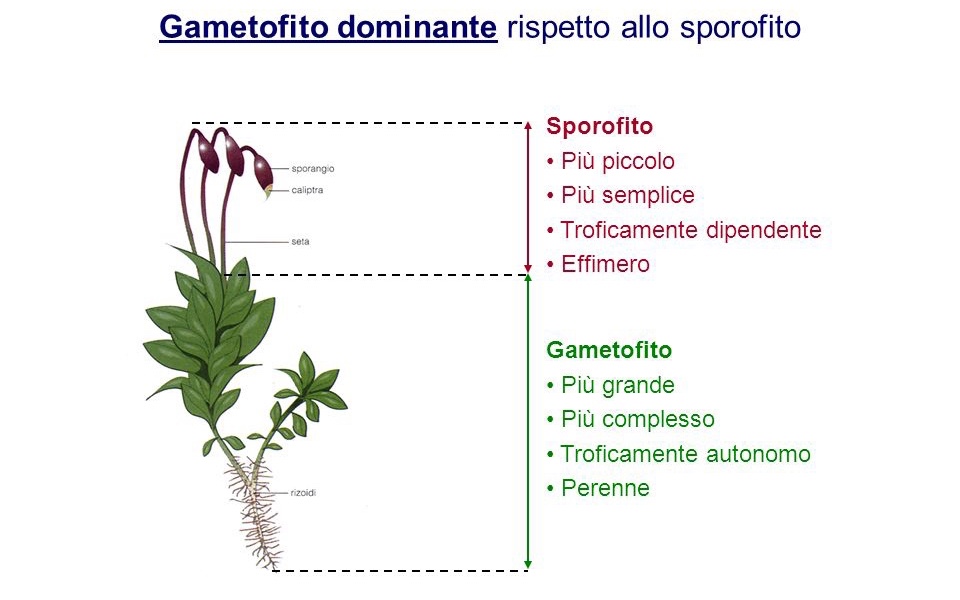
Molte specie sono abbastanza tolleranti ai ripetuti cicli di essiccamento e bagnatura (Clausen 1964, Wood 2007), una caratteristica che ha permesso loro di sfruttare anche i substrati epifitici tra cui foglie e rami. Come le altre briofite, hanno un ciclo vitale eteromorfo con una sporofita dalla vita relativamente breve e dipendente per la nutrizione dal Gametofito, solitamente perenne. Tuttavia, si differenziano da entrambi questi gruppi per numerose caratteristiche citologiche, biochimiche e anatomiche.
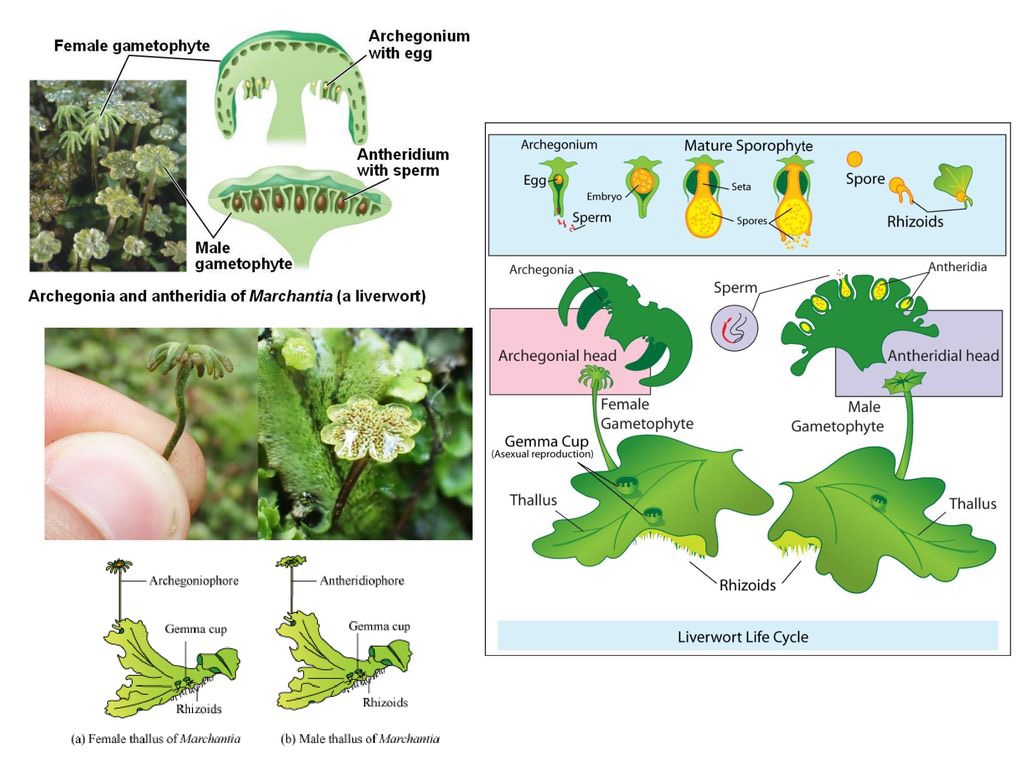
Caratteri diagnostici significativi:
- tendono ad avere un aspetto appiattito anche quando sono frondose, perché le loro foglie sono sempre disposte in file, mai in spirale.
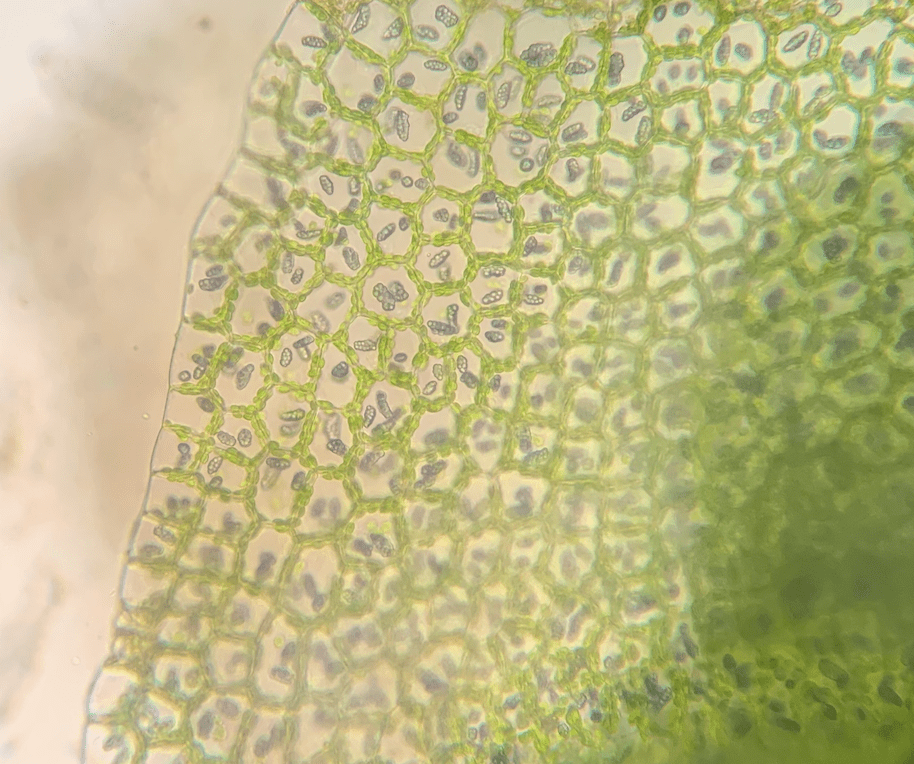
- I rizoidi sono unicellulari, a parete sottile e di solito ialini.
- sia le forme fogliacee che quelle talloidi sviluppano spesso associazioni endosimbiotiche con i funghi;
- gli sporofiti maturano completamente racchiusi dal tessuto gametofitico e sono incapaci di autosostenersi con la fotosintesi;
- le setae dello sporofito sono parenchimatose e si allungano per espansione cellulare piuttosto che per divisione cellulare;
- le capsule sono prive di stomi, cuticola e la columella che sono comuni nei muschi e negli antocerote.




Filogenesi e Morfologia
Le epatiche occupano una posizione critica nell’evoluzione delle piante terrestri, in quanto costituiscono il gruppo gemello di tutte le altre piante terrestri esistenti.
Tradizionalmente, le epatiche sono state suddivise nel gruppo Marchantioide e il gruppo Jungermannioide, che comprende due sottogruppi morfologici, i talloidi semplici anacrogini e i talloidi fogliosi acrogini.
Questi gruppi sono stati definiti nella gerarchia della maggior parte degli schemi di classificazione e sono stati a lungo considerati come unità filogenetiche naturali. Ad esempio, in Crandall-Stotler & Stotler (2000) sono riconosciuti come classi, Marchantiopsida e Jungermanniopsida, con quest’ultima che comprende due sottoclassi, Metzgeriidae (talloidi semplici) e Jungermanniidae (epatiche frondose). Un’ampia serie di caratteri anatomici e di caratteri ontogenetici che differenziano le due classi, tra cui i differenti modelli di sviluppo gametangiale, l’architettura degli spermatidi, l’anatomia della parete della capsula (Crandall-Stotler & Stotler 2000) e i meccanismi coinvolti nella definizione dei piani citochinetici durante la meiosi (Shimamura et al. 2004, Brown & Lemmon 2006). Recenti studi di filogenetica molecolare (per esempio Heinrichs et al. 2005, 2007, Forrest et al. 2006, He-Nygre´n et al. 2006, Qiu et al. 2006) hanno modificato notevolmente questo concetto basato sulla morfologia, soprattutto per quanto riguarda il gruppo dei talloidi semplici.

Corpi oleosi
Le epatiche si distinguono da tutte le altre embriofite per la produzione quasi universale di corpi oleosi, organelli unici legati alla membrana che sintetizzano e sequestrano una vasta gamma di terpenoidi e altri composti aromatici (Flegel & Becker 2000, Suire et al. 2000). I corpi oleosi si formano durante le fasi iniziali della maturazione cellulare (Crandall-Stotler 1981) come dilatazioni del reticolo endoplasmatico (Duckett & Ligrone 1995, Suire 2000) o dalla fusione di vescicole del ditiosoma (Galatis et al. 1978, Apostolakos & Galatis 1998). La membrana che racchiude il corpo dell’olio assomiglia al tonoplasto in quanto ha un aspetto asimmetrico e tripartito, ma si differenzia da esso per la composizione enzimatica e le capacità di trasporto (Suire 2000). L’interno del corpo dell’olio è costituito da piccole goccioline osmiofile sospese in uno stroma granulare ricco di proteine e carboidrati (Pihakaski 1972, Suire 2000).

Calypogeia azurea, b. Cheilolejeunea anthocarpa, c. Radula constricta, d.
Solenostoma truncatum, e. Bazzania tridens, f. Trocholejeunea sandvicensis
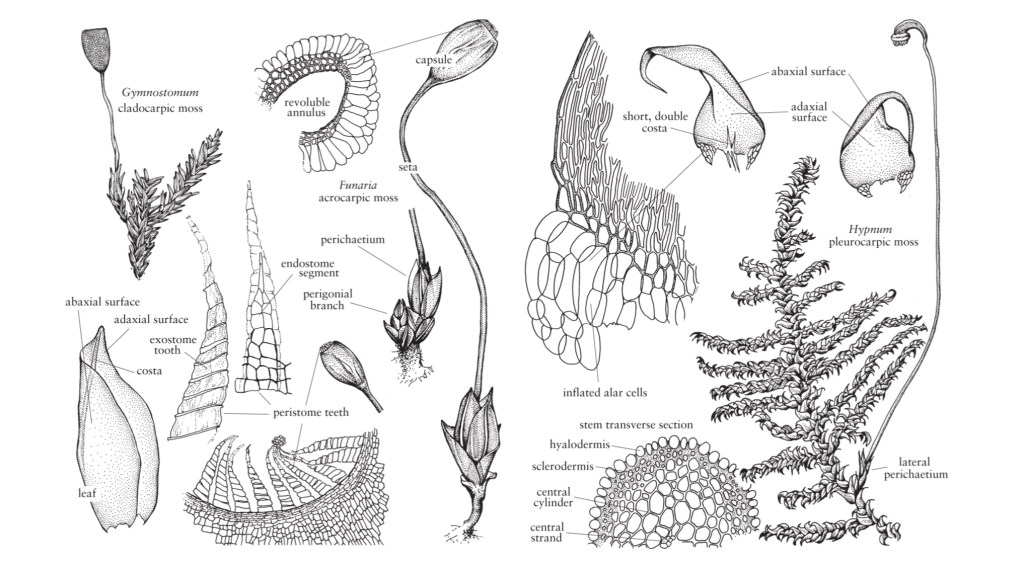
I muschi (Bryophyta)

Con circa 13.000 specie, le Bryophyta costituiscono il secondo phylum più diversificato di piante terrestri.
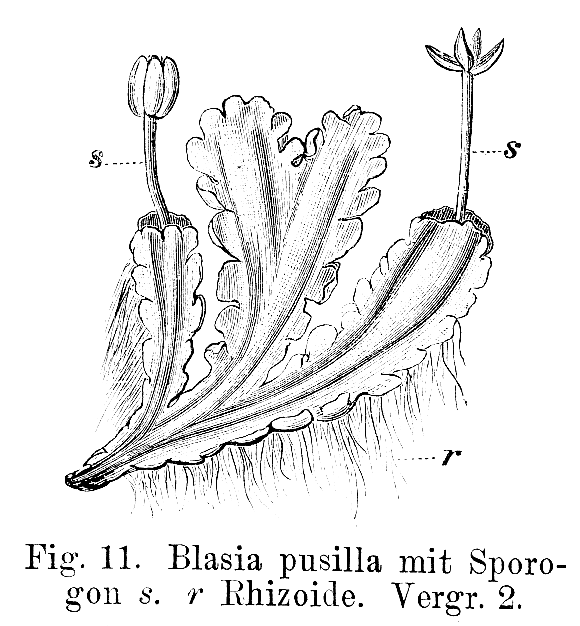
I muschi condividono con le Marchantiophyta e Anthocerotophyta un ciclo vitale aplodiplonte che segna il passaggio dal ciclo vitale dominato dagli aploidi degli antenati delle alghe, al ciclo vitale delle piante vascolari, dominato dagli sporofiti. Il gametofito è libero di muoversi, autotrofo e quasi sempre composto da un fusto frondoso. In seguito alla fecondazione, lo sporofito si sviluppa in un asse non ramificato che porta una capsula terminale portatrice di spore. Lo sporofito rimane fisicamente attaccato al gametofito ed è almeno in parte fisiologicamente dipendente dalla pianta madre.
Recenti ricostruzioni filogenetiche suggeriscono che tre lignaggi delle prime piante terrestri compongano un grado evolutivo che abbraccia il passaggio alla terraferma e l’origine delle piante con sporofiti ramificati. Le Bryophyta sembrano occupare una posizione intermedia: la loro origine precede la divergenza dell’antenato delle antocerote e delle piante vascolari, ma si sono evolute da un antenato comune con le epatiche (Qiu et al. 2006).
L’origine delle prime piante terrestri può essere fatta risalire all’Ordoviciano e forse al Cambriano (Strother et al. 2004). Sebbene fossili inequivocabili di muschi sono stati recuperati solo da sedimenti risalenti a periodi geologici più giovani (Carbonifero superiore), le stime del tempo di divergenza basate sulle filogenesi molecolari suggeriscono che l’origine dei muschi risalga all’Ordoviciano (Newton et al. 2007) e quindi che la loro storia evolutiva, unica nel suo genere, abbracci almeno 400 milioni di anni.
Caratteri del gametofito
Germinazione delle spore
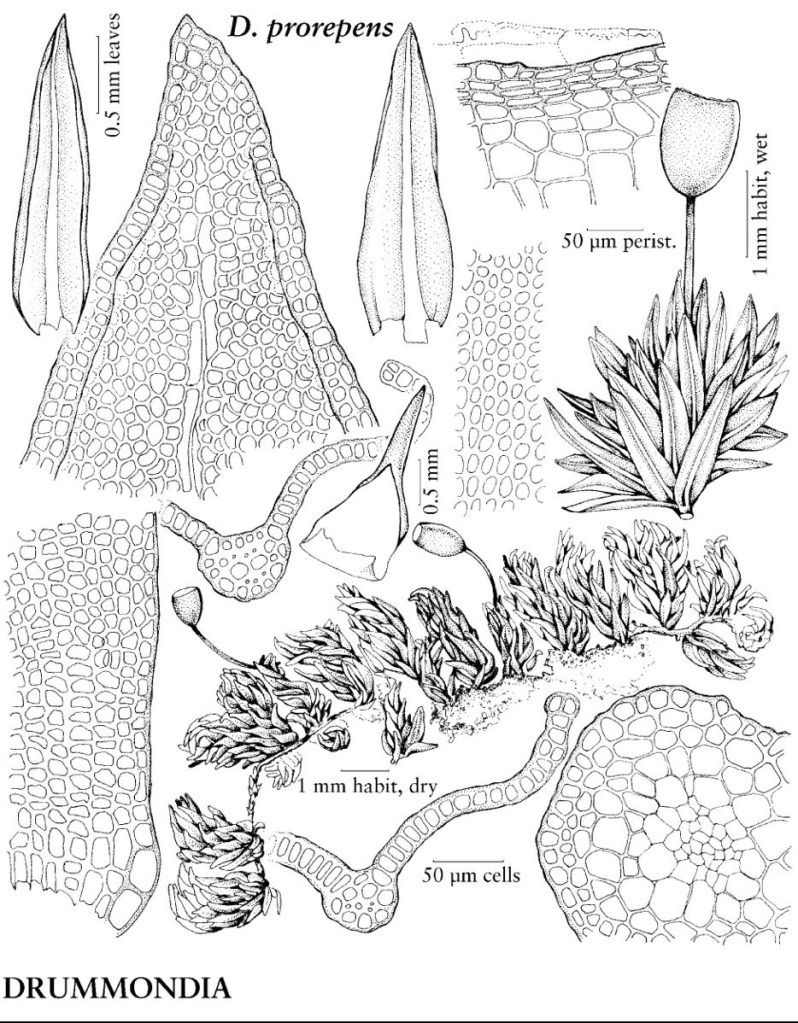
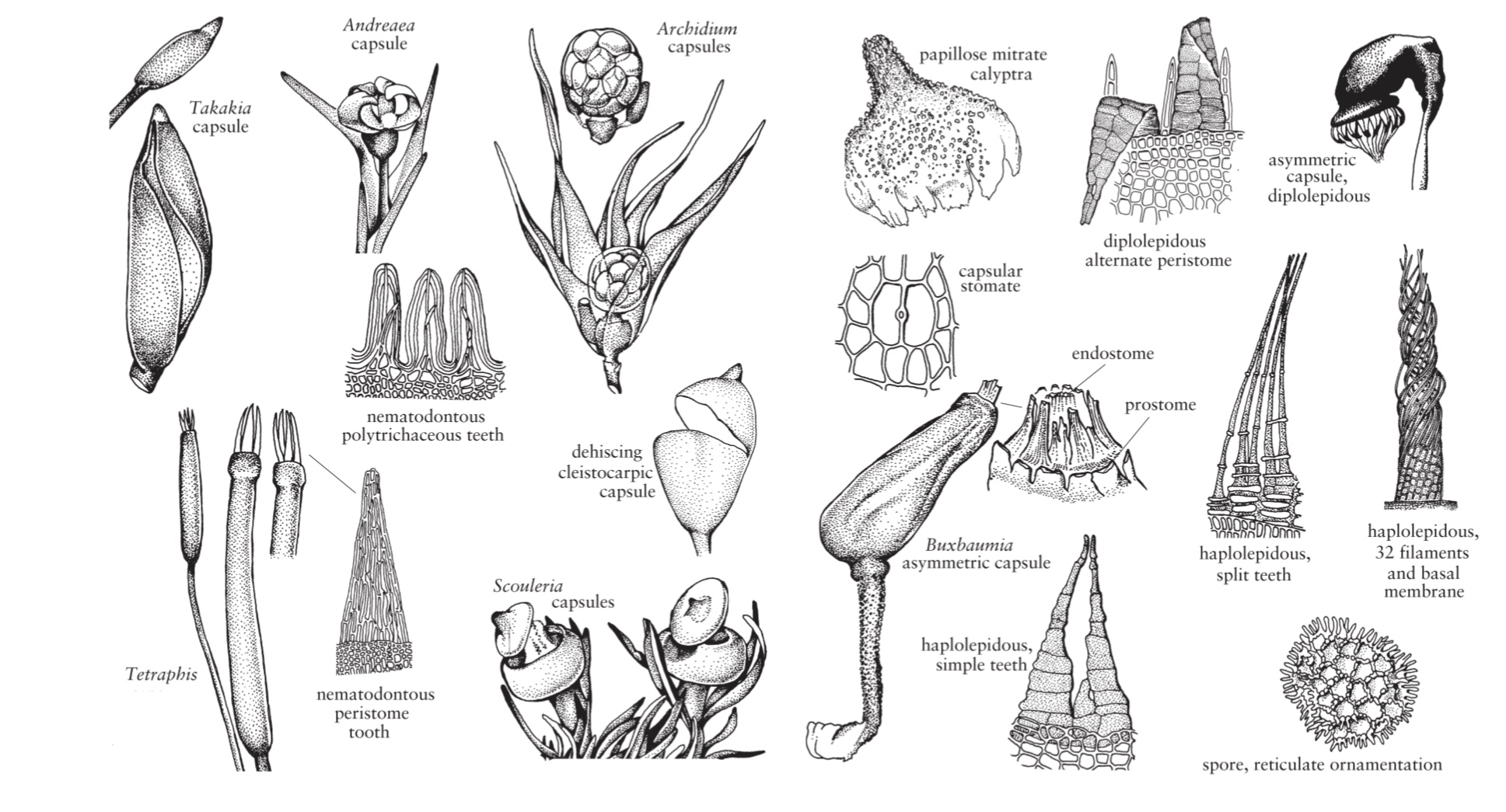
Un muschio inizia il suo ciclo di vita quando le spore aploidi vengono rilasciate da una capsula sporofita e iniziano a germinare. Nella maggior parte dei muschi, la germinazione è esosporica, cioè la parete della spora viene rotta dal protoplasto della spora che si espande prima di qualsiasi divisione cellulare. Tuttavia, in alcuni muschi, ad es, Andreaea, Drummondia e Leucodon, la germinazione è precoce ed endosporica, vale a dire che le divisioni cellulari si verificano prima del rilascio delle spore e della rottura della parete spermatica, rispettivamente. Questi taxa sono descritti come spore multicellulari.

La specializzazione cellulare si verifica all’interno del protonema per formare due tipi di filamenti, un sistema orizzontale di filamenti di ancoraggio di colore marrone rossastro, chiamato caulonema, e filamenti eretti di colore verde, il cloronema. Ogni protonema può estendersi per diversi centimetri, formando una pellicola verde sul suo substrato.
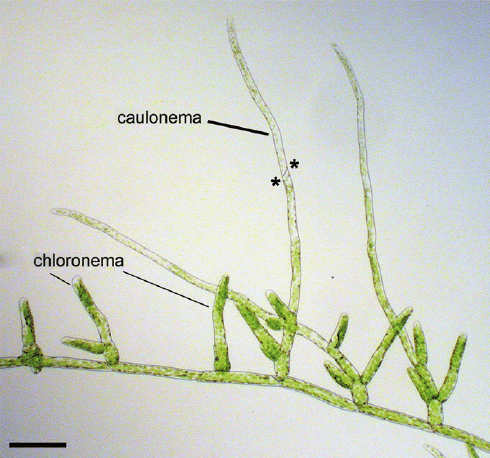
Frammenti di cloronema, tagliati dalla formazione di cellule specializzate per l’abscissione (tmema), possono disperdere ulteriormente il protonema. Man mano che il protonema cresce, le cellule bersaglio, di solito sul caulonema generano iniziali di gemme che si divideranno infine con divisioni oblique sequenziali per formare le cellule apicali della gemma. Questo dà inizio alla crescita del gametoforo fogliare o germoglio del muschio.
Morfologia e abitudini dei germogli
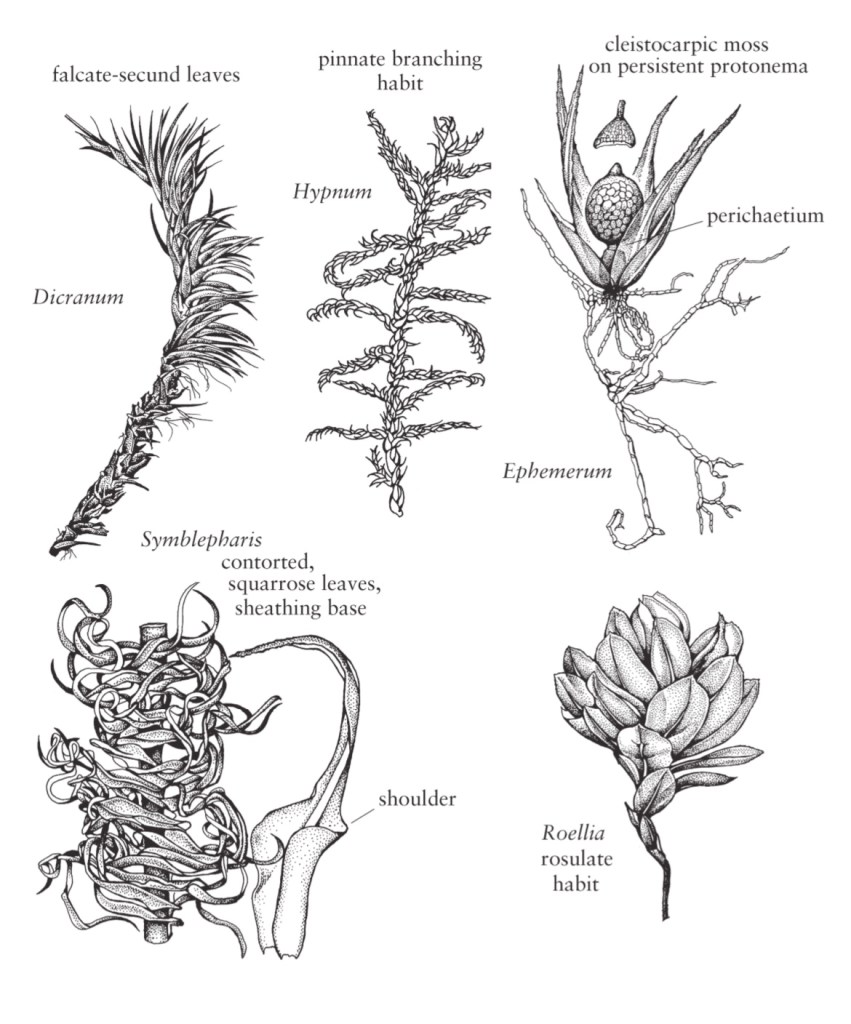
Il germoglio frondoso continua a crescere grazie all’attività mitotica della sua cellula apicale da obovoidale a fusiforme e del meristema circostante. Le divisioni che si verificano nella cellula apicale formano derivati disposti a spirale, ognuno dei quali darà origine a una singola foglia e una porzione del fusto. L’angolo di divergenza tra i derivati successivi è responsabile della disposizione spaziale delle foglie o della fillotassi del germoglio.

In alcuni muschi, le foglie mature sono chiaramente disposte su due file, ad es. Fissidens e Bryoxiphium, una fillotassi di 1/2, mentre Fontinalis e Tetraphis hanno foglie allineate su tre file, una fillotassi di 1/3. La maggior parte dei muschi, tuttavia, ha foglie distribuite a spirale, con le fillotassi 2/5 e 3/8 che sono le più comuni (W. Frey 1971; B. Crandall-Stotler 1984).

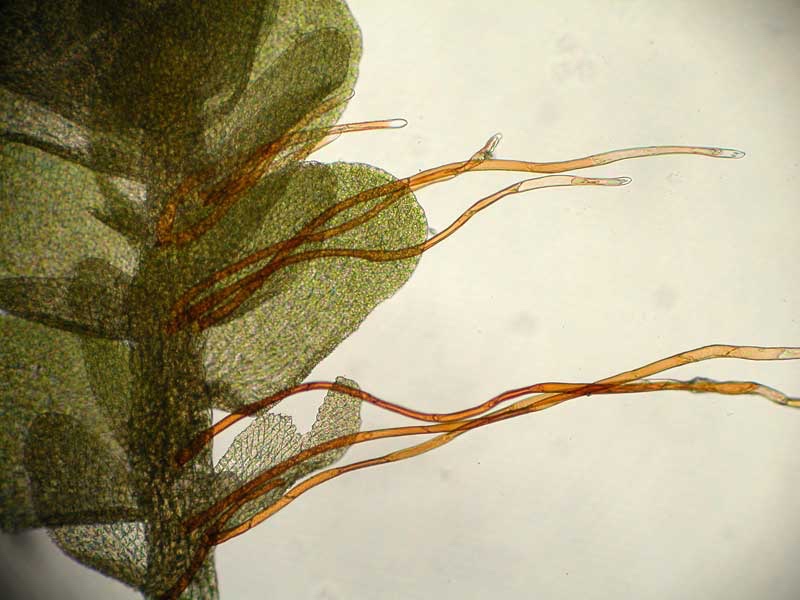
Rizoidi
Ad eccezione della Takakia e dello Sphagnum, i muschi sono ancorati al loro substrato da rizoidi filamentosi, spesso ramificati, di colore marrone rossastro. Come nei caulonemata, i rizoidi sono
multicellulari con pareti trasversali oblique; le loro pareti sono lisce o ruvide con papille. I rizoidi possono essere alla base del fusto, soprattutto negli acrocarpi, o sorgono lungo tutto il fusto a contatto con il substrato. In alcuni taxa, come ad esempio Tomentypnum, i rizoidi densamente impacchettati formano un tomento simile al feltro sul fusto.
Fotografia: Heino Lepp
La maggior parte dei rizoidi sono sottili e poco ramificati (tipo micronematale), ma altri sono di diametro maggiore e ampiamente ramificati (tipo macronematale). I rizoidi non sono i principali siti di assorbimento di acqua e nutrienti, ma possono favorire il movimento capillare dell’acqua lungo la superficie esterna del fusto (M. C. F. Proctor 1984). Funzionano principalmente come strutture di ancoraggio e in alcuni taxa, come Leptobryum e Pyramidula, formano dei propaguli asessuati che aiutano la diffusione della colonia su substrati non occupati.
Foglie
La notevole variazione che si verifica nella disposizione e la struttura delle foglie di muschio fornisce alcuni dei caratteri più utili nell’identificazione della specie. Le foglie nascono tipicamente da tutti i lati del fusto, la maggior parte delle quali a spirale, ma si possono trovare anche disposizioni distiche e tristiche. Le foglie mature di un determinato germoglio sono di solito tutte simili in forma e dimensione, cioè isofille, ma esistono taxa che sono anisofillie, con foglie dorsali o ventrali decisamente più piccole di quelle laterali.
Anatomia del fusto
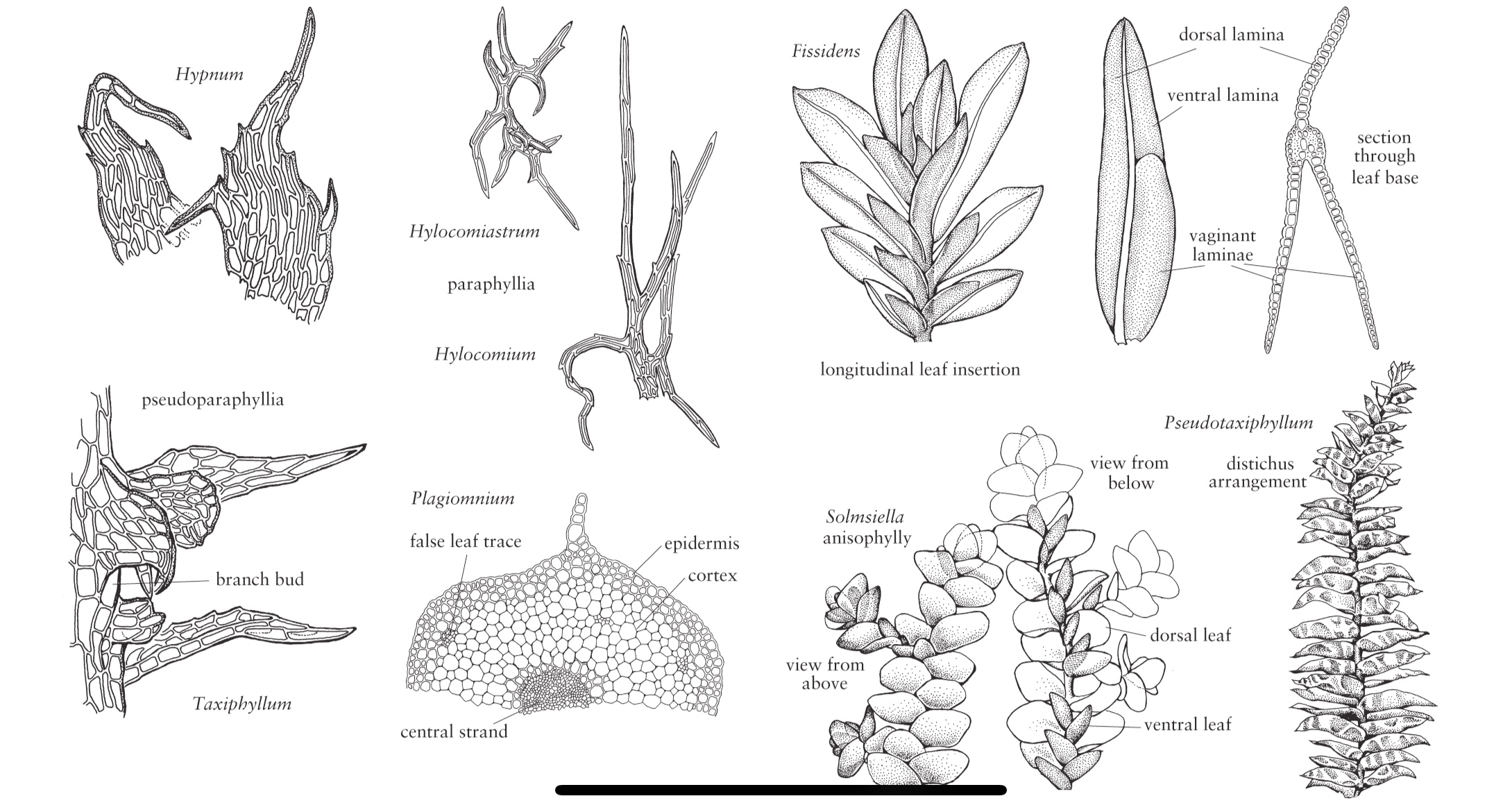
In molti muschi, il fusto è anatomicamente complesso, costituito da uno strato epidermico differenziato, da una corteccia e da un filamento centrale di cellule idrolizzate conduttrici d’acqua a parete sottile, chiamate idroidi.
Caratteri delle sporofite
Sviluppo dell’embrione
Concomitante con la crescita dello sporofito embrionale, divisioni cellulari nel centro archegoniale circostante, nel peduncolo archegoniale basale e nel germoglio del gametofito sotteso producono un epigonio che lo racchiude. All’inizio dello sviluppo, una zona di trasferimento di sostanze nutritive, o placenta, si differenzia tra il piede conico dello sporofito e la parte basale dell’epigonio. Sia i nutrienti organici che l’acqua si spostano dal gametofito allo sporofito attraverso la placenta. Al di sopra del piede si differenzia una seta simile a uno stelo, le iniziali della capsula embrionale vengono staccate da una cellula apicale all’apice dello sporofito. La creazione di un meristema della seta appena sotto la capsula in via di differenziazione allunga lo seta, strappando l’epigonio. La parte basale dell’epigonio, che racchiude ancora la parte inferiore dello seta e del piede, viene ora chiamato vaginula e la parte superiore che rimane sopra la punta dello sporofito è la calyptra.
Anatomia della seta
La seta e la capsula continuano a svilupparsi dopo l’emersione dello sporòfito dall’epigonio, la seta da un meristema apicale generalizzato e la capsula da divisioni modellate nelle iniziali della capsula precedentemente formate.
Di solito matura un solo sporofito per ogni gineceo, ma in alcuni taxa, ad esempio Dicranum, se ne può sviluppare più di uno, risultante una polisetia, ovvero più setae che emergono da un singolo perichezio.
Dal punto di vista anatomico, la seta assomiglia allo stelo del gametofito. Prima della maturazione della capsula, la seta è tipicamente clorofillosa, ma a maturità diventa rossastro o bruno-giallastro, a causa di pigmenti depositati nelle pareti ispessite delle cellule epidermiche e degli stereidi. Spesso le file di cellule epidermiche sono allineate in modo elicoidale; questa disposizione, così come il diverso spessore della parete sui diversi lati permette alla seta di attorcigliarsi quando si asciuga, e di distorcersi quando si reidrata.
Anatomia della capsula
I muschi sono monosporangiati, cioè ogni sporofito produce un solo sporangio o capsula.
Le variazioni nella morfologia della capsula matura forniscono una ricchezza di caratteri sistematicamente importanti. La forma delle capsule varia da sferoide a ovoidale, obovoide, piriforme, turbinata, ellissoide o lungo-cilindrica, e possono essere simmetrici, come Orthotrichum, o asimmetrici, come ad esempio nella Funaria. Possono essere erette, inclinate, annuenti (cernuote) o addirittura pendenti. L’esotecio, o epidermide della capsula reca uno strato lucido simile a una cuticola. Le capsule immature sono verdi, ma a maturità sono di varie tonalità di giallo, rosso o marrone.
Tipi distinti di anatomia della capsula caratterizzano i cinque gruppi principali o superclassi di muschi. Nella maggioranza dei muschi veri (Superclasse V), le capsule sono costituite da tre zone anatomicamente distinte, vale a dire il collo basale, l’urna o teca mediana contenente le spore e l’opercolo distale.
Le capsule con opercolo deiscente sono stegocarpiche. Le capsule prive di opercolo e che presentano deiscenza irregolare a causa di una rottura generalizzata della pareti della capsula sono cleistocarpe.


Lascia un commento